저널의 신경학,신경과학
키워드
SID;Me5;해마다,수면 단계;SIUDS;젖꼭지;SUDC
소개
젖꼭지 사용 및 아닌 영양 빨
아닌 영양 흡입은 정상적인 태아와 신생아는 동작입니다. 젖꼭지(인형이라고도 함)는 2000 년 이상이 타고난 욕구를 충족시키는 데 사용되었습니다. 이 모의 젖꼭지는 유아를 진정 시키거나 진정시킬 수 있으며 사소한 의료 절차 중에도 유용합니다. 예를 들어,젖꼭지 빨기는 venipuncture 를 겪고있는 어린 영아에서 울음을 감소시키는 것으로 나타났습니다. 1979 년에 젖꼭지 사용이 sids 가 수면 무호흡증과 관련이있을 때 갑자기 유아 사망 증후군(sids)의 위험을 감소시킬 수 있다고 가정했습니다. 이 관계를 증명되지도 않기 때문가 강하고 일관적 증거는 더 적은 유아 SID 젖꼭지를 사용하는 동안 밤에 잠을 보다는 나이 일치 제어를 제공합니다. 고무 젖꼭지가 SIDS 에 대한 보호를 제공하는 메커니즘은 두 가지 메커니즘이 제안되었지만 불분명합니다. 첫째,보호는 자율 통제에 미치는 영향과 관련이있을 수 있습니다. 둘째,젖꼭지는 수면에서 각성을 증가시킬 수 있습니다.
Me5
의 목적은 우리의 연구는 mesencephalic 삼차 핵(Me5),독특한 신경 형성에 중추 신경계(CNS)으로만 사이트의 내 neuraxial 신경절에 있는. 즉,일차 구 심성 감각 뉴런의 세포체를 포함합니다. 이 좁은 세포 밴드는 periaqueductal gray(PAG)바로 옆을 지나고 pons Varolii 와 midbrain 사이의 경계에서부터 우수한 colliculus(midbrain 의 상한)까지 확장됩니다. 그것의 하한은 모터 삼차 핵(Mo5)에 rostral 입니다. Me5 옆에는 더 내측으로 그리고 네 번째 뇌실 앞에는 신경계의 노르 아드레날린 성 섬유의 주요 공급원 인 locus coeruleus 가 있습니다. LC 와 Me5 는 초기 발달에 밀접하게 관련되어있다;LC 는 적절한 LC 기능에 필요한 Me5 뉴런의 분화에 중요합니다.
Me5 은 완전히 둘러싸여 있으로 형성(RF),그리고 그것의 꼬리 부분은 핵심으로 구성된 작은,다극,부르는 gamma-aminobutyric acid(GABA)ergic 세포 rostral 을 Mo5 고 우리는 나타내로 Me5c(꼬리 부분의 mesencephalic 삼차 핵). 반대로,rostral 부분은 midbrain 에 앉고 수상 돌기가없는 큰 의사 단극 glutamatergic 세포로 구성됩니다.
Me5 세포는 소그룹의 뉴런 사이에서 소마토-체세포 시냅스를 형성하는 갭 접합부를 갖는다. 각 신경은 거의 완전히 덮여 프로세스에서 방출 하는 두 개 또는 더 이다(일반적으로 이다 더 많은 신경). 중뇌 삼차 핵 뉴런의 말초 가지는 주로 치주 인대의 기계 수용체를 내재화한다. 이러한 수용체는 매우 민감한 운동 엘리베이터의 근육에서는 턱;그들에 의해 활성화된 간단한 터치로의 치아를 매우 작은 힘이 필요하(1N 및 4N 에 전치 및 사후 치아,respectively). 또한,Me5may 의 치주 기계 수용체는 자극이없는 상태에서”발사”합니다. 중앙 가지는 모터 삼차 핵,망상 parvocellular 지역 및 dorsolateral midbrain RF(tegmentum 의 등쪽 raphe 핵 및 측 방향 등쪽 핵)로 이동합니다. 때 그들을 통해 내려 Varolio 의 다리로”스 Probst,”그들은 십자가 dorsolateral 구의 수준에서 등의 핵 미주신경로 전달 측면 피개 필드를 종료하고 척수에서 수준에서의 꼬리가 삼차 핵 최대 C1/C3. 독방에 복부에 위치한 핵 Probst 의 큰 뉴런을 통과하면 hypoglossal nucleus,trigeminal main sensory 및 caudate nucleus,독방 핵,그리고 결국 supra-trigeminal nucleus 에 도달합니다. 의 신경 세포 midbrain RF,특히 이들의 drn 에와 LDT,모두가 직접을 통해 시상에 보내는 광범위한 특정 계획을 전체 피질과 양식의 일부분을 오름차순으로 활성화 시스템(ARAS). Me5 는 큰 의사 단극 뉴런과 작은 Me5c 뉴런(그림 1)으로부터의 GABAergic 시그널링을 통해 Mo5 에 glutamatergic 시그널링을 제공합니다.
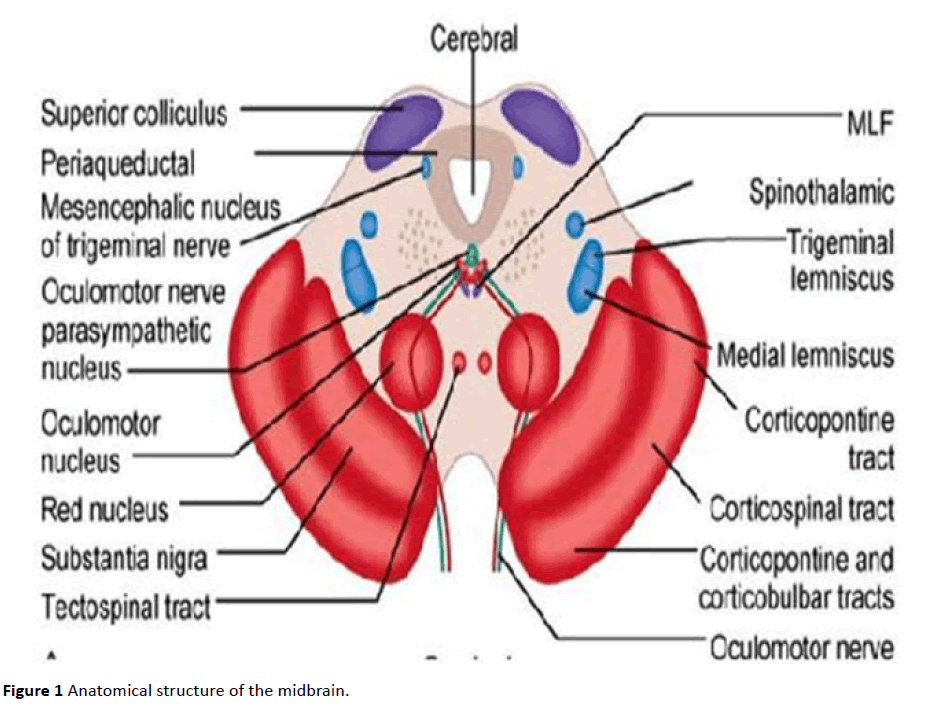
그림 1 중뇌의 해부학 적 구조.
유아 수면 패턴
전과 후 즉시 출생,수면이 본질적으로만 rapid eye movement(REM)sleep,하지만 시간의 비 REM(NREM)수면 급속하게 증가한 첫 달에서의 삶입니다. 실제로,유아 수면 더 정확한 것으로”행동 상태”라고 조용한 수면(QS 에 해당하는 NREM sleep),활성 수면(로에 해당하는 REM sleep),무한한 잠(IS),그리고 깨어(W,활성 또는 자). 신생아를 구분하지 않습은 낮과 밤의 25 시간 신생아 수면에 독립적인 환경 및 규율에 의해서만이 내부 필요 같은 배고픔과 갈증을.
수면은 시상 하부의 VLPO 와 MNPO 가 뇌간 표적,특히 ARAS(DRN,LC,TMN 등)에서 GABA 와 galanin 을 전달할 때 발생합니다.). 아세틸콜린,도파민,히스타민,세 orexin 생산의 감소,고가의 억제 부교감 핵 LC 제공하는 노르에피네프린해 교감 신경절 척추 신경.
가설
아기가 잠들기 시작하면 VLPO 와 MNPO 핵이 gaba 를 뇌간과 시상 하부로 방출해야합니다.
gaba 가 표적 세포에 도달하면 막 전위차가 염화물 진입에 따라 매우 음성이기 때문에 억제됩니다. 이 저해 방지 신경전달물질의 방출 및 활동의 중요한 단백질(예:도파민은 수송이나 모노 산화효소). 따라서 SID 를 가진 유아의 부검 소견은 이들 단백질의 결핍 수준을 나타낼 가능성이 높습니다. 그들의 결핍은 원인보다는 SIDS 의 영향 일 가능성이 높습니다. 수면 중 특정 시점에서 GABA 수준이 증가하고 많은 세포가 억제되면 뇌간의 본질적인 메커니즘이 활성화됩니다. Me5 는 주로 의사 단극 세포로 구성되어 있지만 꼬리 종단(Me5c)은 일반적으로 GABAergic 인 작은 다극 세포로 구성됩니다. 그들은 Mo5 에서 바로 맞은 편에 위치하고 있기 때문에,그들은 일반적으로 휴식 조건 하에서 억제됩니다. 시상 하부에서 방출 된 GABA 가 Me5c 세포를 억제하면 Mo5 의 Me5 억제가 해제됩니다. 저작 근육은 수축하고 치아가 닿아 Me5 를 활성화시키고 글루타메이트를 ARAS 핵,PAG 및 부교감 핵에 방출하게합니다. 순 결과는 GABA 의 억제 효과의 감쇠입니다. 이러한 조건에서,특정 신경 세포가 죽고 활성화 교 셀 릴리스 IL1 베타 프로스타글란딘는 늘 물질 P 수준이다. 이러한 사건은 빈번하지만 치명적인 결과로 끝나는 경우는 거의 없습니다; 그러나,그들은 영향을 미칠 수 있습 유아 발달을 극도의 약점”brainstem 시스템,”많은 핵세포는 제대로 작동하지 않습니다. 에서 이 시나리오,아이 높은”감도”수많은 요소는 일반적으로 치명적이 아닌나가 될 수 있도의 약점으로 인해 시스템에서는 시간(예:긴 QT,사소한 감염,etc.).
이러한 맥락에서 우리는 gliosis;leukomalacia;뇌 저형성;증가 된 물질 P 수준; 세로토닌,도파민,아세틸 콜린,노르 에피네프린,히스타민 및 오렉신 수준의 감소.
중요한 요소 중 하나는 수면 위치입니다. 경향이있는 위치에서 모든 호흡은 척추,폐 및 심장을 포함하는 흉곽을 확대하기 위해 더 큰 작업이 필요합니다. 신생아에서 이러한 구조물의 무게는 3~4kg 의 총 무게로~0.5kg 입니다. 그것은 흉곽에~10kg 의 체중으로 등 뒤에서 자고있는 80kg 의 성인 남성과 비슷합니다. 호흡기 시스템의 체중이 특정 신경 전달 물질,특히 세로토닌에 의해지지되지 않을 때 사망이 발생할 수 있습니다. 젖꼭지는 Me5 를 활성화 할 수 있으며,그 다음 글루타메이트를 ARAS 핵과 그 표적에 방출하여 과도한 GABAergic 전달 및 뇌간”약점을 예방합니다.”
결론
우리의 etio-pathogenesis 모델은 SIDS 의 특성과 일치하는 많은 양상을 포함한다. 특히 젖꼭지 사용이 SIDS 를 예방할 수있는 이유를 설명합니다. 또한 수면 중에 신경 전달 불균형이 특히 위험한 이유를 강조합니다. 주목할 만하게,그것은 SIDS 를 가진 유아의 해마에있는 부검 소견의 대부분을 설명 할 수있다. 이것은 Me5 섬유의 양측 병변이 유사한 해마 상아질 이랑 손상으로 이끄는 동물 연구에 의해 뒷받침된다.
우리는 이상에서 찾을 SID 뇌하지 않은 결과의 친밀한 관계 Me5 과 해마다;오히려,그들은 그로 인해 변경 REM sleep 에 의해 발생 Me5 능. 에서 출생,신경에 응답해야 합니다 그들의 필요를 통해 전체의 표현이 적절한 유전자,그러나 이 시스템은 실패할 수 있습니다,특히에서의 컨텍스트 환경 문제와 같은 담배 또는 자고 쉬운 위치에. 이러한 영향은 방출 된 이산화탄소를 재 도입하거나 리브 케이지 확장에 도전함으로써 호흡을보다 어렵고 덜 효율적으로 만듭니다. 이것은 중요한 중뇌 구조의 성숙을 막고 SIDS 로 이어질 수 있습니다.
이해의 충돌
저자가 선언하는 연구가 수행되었의 부재에서 어떠한 상업 또는 금융 서비스를 제공하는 것으로 해석될 수 있는 잠재적인 이해의 충돌.
윤리 선언문
원고에 제시된 연구는 인간 또는 동물 과목을 포함하지 않습니다.
- 레빈 S(1971)인형. S Afr 의대 J45:237-240.
- Zempsky WT,Cravero JP(2004)American Academy Of Pediatrics Committee On Pediatric Emergency Medicine And Section on Anesthesiology And Pain Medicine. 응급 의료 시스템에서 소아 환자의 통증 및 불안 완화. 소아과 114:1348-1356.
- Curtis SJ,Jou H,Ali S,Vandermeer B,Klassen T(2007)소아 응급실에서 venipuncture 를받는 영아를위한 진통제로서 자당 및/또는 젖꼭지의 무작위 대조 시험. BMC 소아과 7:27.
- Cozzi F,Albani R,Cardi E(1979)갑작스런 침대 사망 및 수면 무호흡증에 대한 일반적인 patho-생리학. ‘진공-글로소-안검 하수증 증후군’. MedHypotheses5:329-338.
- Yiallourou SR,Poole H,Prathivadi P,Odoi A,Wong FY,et al. (2014)수면 중 유아 혈압 및 자율 활동에 대한 더미/젖꼭지 사용의 효과. 수면 의대 15:1508-1516.
- Franco P,Scaillet S,Wermenbol V,Valente F,Groswasser J,외. (2000)수면에서 유아의 각성에 대한 젖꼭지의 영향. J 소아과 136:775-779.
- Hanzer M,Zotter H,Sauseng W,Pfurtscheller K,Müller W,외. (2009)젖꼭지 사용은 수면중인 유아의 자발적인 각성의 빈도 또는 지속 시간을 변경하지 않습니다. 수면 의대 10:464-470.
- Espana A,Clotman F(2012)One cut factors 는 locus coeruleus 와 mesencephalic trigeminal nucleus 의 발달을 제어합니다. 몰 세포 신경 과학 50:93-102.
- Morgane PJ,Jacobs MS(1979)raphe 는 쥐의 locus coeruleus 에 대한 투영. 뇌 고해상도 황소 4:519-534.
- Hayar A,Poulter mo,Pelkey K,Feltz P,Marshall KC(1997)감마-아미노 부티르산에 대한 중뇌 삼차 뉴런 반응. 뇌 해상도 753:120-127.
- Copray JC,Liem RS(1993)생체외에서 래트의 중뇌 삼차 뉴런의 생존 및 뉴라이트 형성. 아치 구강 바이올 38:547-557.저작 근육의 양측 신경 분포에 대한
- Rokx JT,Juch PJ,Van Willigen JD(1985):역행 추적자를 사용한 연구. J Anat140:237-243.
- 백 SK,곽 MK,안 DK,김 YK,김 DS 등. (2000)쥐 삼차 중뇌 핵 내의 턱 근육 스핀들 구 심성의 초 구조. 뉴로 포트 16:1561-1564.
- lazarov NE(2000)고양이의 중뇌 삼차 핵. Adv Anat Embryol 세포 Biol153:1-103.
- Henderson G,Pepper CM,Shefner SA(1982)체외에서 삼차 신경의 국소 coeruleus 및 mesencephalic 핵에 포함 된 뉴런의 전기 생리 학적 특성. Exp Brain Res45:29-37.
- Curti S,Hoge G,Nagy JI,Pereda AE(2012)전기 결합과 막 특성 사이의 시너지 효과는 중뇌 삼차 핵의 뉴런의 강력한 동기화를 촉진합니다. J Neurosci32:4341-4359.
- Liem RS,Copray JC,Van Willigen JD(1991)쥐 중뇌 삼차 핵의 초 구조. Acta Anat(바젤)140:112-119.
- Trulsson M(2006)인간 치주 기계 수용체의 감각 운동 기능. J 구두 Rehabil33:262-273.
- Trulsson M(2007)유양 돌기 동안 인간 치주 기계 수용체에 의한 강제 인코딩. 아치 구강 바이올 52:357-360.
- Trulsson M,Gunne HSJ(1998)치주 수용체가 결여 된 인간 피험자에서 음식 보유 및 물기 행동. J 덴트 고해상도 77:574-582.
- Trulsson M,Johansson RS,Olsson KA(1992)치아에 가해지는 힘에 대한 인간 치주 mechanoreceptive afferents 의 방향 감도. J Physiol447:373-389.
- Rokx JT,Juch PJ,Van Willigen JD(1986)래트에서 중뇌 삼차 뉴런의 배열 및 연결. 액타 아낫(바젤)127:7-15.
- Matsushita M,Okado N,Ikeda M,Hosoya Y(1981)는 삼차 신경의 척수 및 중뇌 핵으로부터 고양이의 척수로의 하강 투영. 양 고추 냉이 퍼 옥시 다제 기술을 사용한 연구. J Comp Neurol196:173-187.
- Sirkin DW,Feng AS(1987)쥐의 폰틴 망상 형성 및 중뇌 삼차 핵으로부터의 하강 경로의 자동 방사선 학적 연구. J Comp Neurol256:483-493.
- Nieuwenhuys R,Voogd JD,Van Huijzen C(2008)인간 중추 신경계. 뉴욕:스프링거.
- Matesz C(1981)래트에서 중뇌 삼차 뿌리의 섬유의 말초 및 중심 분포. 신경 과학 레트 27:13-17.
- Kolta A,Westberg KG,Lund JP(2000)토끼의 삼차 운동 핵과 인접한 구조에 투영되는 뇌간 내측의 식별. J Chem Neuroanat19:175-195.
- Hayar A,Poulter mo,Pelkey K,Feltz P,Marshall KC(1997)감마-아미노 부티르산에 대한 중뇌 삼차 뉴런 반응. 뇌 해상도 753:120-127.
- Meier-Koll A(1979)출생 후 발달 동안 내인성 리듬의 상호 작용. 한 명의 정상적인 유아에서의 행동 및 거짓말 탐지기 연구의 관찰. Int J Chronobiol6:179-189.
- Kinney HC,Cryan JB,Haynes RL,Paterson DS,Haas EA,et al. (2015)유아에서 갑자기 설명 할 수없는 사망의 상아질 이랑 이상:근본적인 뇌 취약성의 형태 학적 마커. Acta Neuropathol129:65-80.
- 이시이 T,스에나가 R,이와타 W,미야타 R,후지카와 R 등 (2010)중뇌 삼차 감각 핵의 양측 병변은 해마 신경 발생을 자극하지만 공간 기억 재설정에 심각한 적자를 초래합니다. 뇌 해상도 1342:74-84.
- Lopez J,Roffwarg HP,Dreher A,Bissette G,Karolewicz B,et al. (2008)빠른 안구 운동 수면 박탈은 장기 전위 안정성을 감소시키고 해마 발달 동안 일부 글루타메이트 성 신호 단백질에 영향을 미친다. 신경 과학 153:44-53.