解剖学

すべての脊椎動物は、同様の基本的な体硬直棒、脊索;神経材料の背側中空管、神経管;咽頭アーチ;および肛門の後部の尾。 脊髄は脊柱によって保護され、脊索の上にあり、胃腸管はその下にある。 神経組織は外胚葉に由来し、結合組織は中胚葉に由来し、腸は内胚葉に由来する。 後端には脊髄と椎骨を続ける尾がありますが、腸は続きません。 口は動物の前端にあり、肛門は尾の基部にあります。 脊椎動物の決定的な特徴は、脊柱であり、分割された一連の椎骨の発達において形成される。 ほとんどの脊椎動物では、脊索は椎間板の髄核になる。 しかし、チョウザメやシーラカンスのようないくつかの脊椎動物は、成人期に脊索を保持しています。 顎の脊椎動物は二次的に失われるかもしれない対になった付属器、ひれまたは足によって代表されます。 脊椎動物の四肢は、同じ基礎となる骨格構造が彼らの最後の共通の祖先から継承されたので、相同であると考えられている。 これはチャールズ-ダーウィンが進化論を支持するために提唱した議論の一つである。
魚の解剖学編集

魚の体は頭、胴、尾に分かれていますが、三つの間の分割は常に外部に見える 魚の内部の支持構造を形成する骨格は、軟骨、軟骨魚、または骨の魚の骨のいずれかで作られています。 主な骨格要素は、軽量でありながら強い関節椎骨で構成される脊柱である。 肋骨は背骨に付着し、四肢や四肢のガードルはありません。 魚の主な外部の特徴、フィンは、尾鰭を除いて背骨と直接接続していない光線と呼ばれる骨または柔らかい棘のいずれかで構成されています。 それらは、胴体の主要部分を構成する筋肉によって支持されている。 心臓には2つのチャンバーがあり、鰓の呼吸面を通って血液をポンプで送り、単一の循環ループで体の周りに送ります。 目は水中を見るために適応しており、局所的な視力しか持たない。 内耳はありますが、外耳または中耳はありません。 低周波振動は、魚の側面の長さに沿って走る感覚器官の側線システムによって検出され、これらは近くの動きや水圧の変化に応答する。
サメやエイは、軟骨で構成される骨格を含む、古代の魚のものと同様の多数の原始的な解剖学的特徴を持つ基底魚です。 彼らの体は背腹側に平らになる傾向があり、通常は5対の鰓のスリットと頭の下側に大きな口があります。 真皮は、別個の真皮プラコイド鱗屑で覆われている。 彼らは尿と生殖器の通路が開いているクロアカを持っていますが、水泳の膀胱はありません。 軟骨魚は、小さな数の大きな卵を産生する。 いくつかの種は卵胎生であり、若者は内部的に発達するが、他の種は卵胎生であり、幼虫は卵の場合に外部的に発達する。
骨の魚の系統は、多くの場合、古代の魚の特徴からの主要な進化的変化を伴う、より派生した解剖学的特徴を示しています。 彼らは骨の骨格を持ち、一般的に横方向に平らにされ、手術室によって保護された鰓の五対を持ち、鼻の先端またはその近くに口を持っています。 真皮は重なり合う鱗で覆われている。 骨のある魚にそれらが水コラムの一定した深さを維持するのを助けるcloacaない水泳のぼうこうがある。 彼らは主に、彼らは水柱に放送少し卵黄と小さな卵を大量に産卵します。h3>
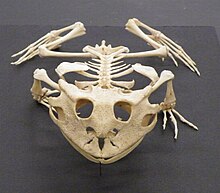
両生類は、カエル、サンショウウオとcaeciliansからなる動物のクラスです。 彼らは四足動物ですが、caeciliansとサンショウウオのいくつかの種は、手足がないか、その手足がはるかにサイズが縮小されているのいずれかを持っています。 彼らの主な骨は中空で軽量であり、完全に骨化されており、椎骨は互いに連動し、関節突起を有する。 彼らの肋骨は通常短く、椎骨に融合することができます。 彼らの頭蓋骨は主に広くて短く、しばしば不完全に骨化されています。 彼らの皮膚にはケラチンがほとんど含まれておらず、鱗が欠けていますが、多くの粘液腺といくつかの種では毒腺が含まれています。 両生類の心臓には3つの室、2つの心房と1つの心室があります。 彼らは膀胱を持っており、窒素性廃棄物は主に尿素として排泄されます。 両生類は、鼻孔を通って口腔咽頭領域に空気が最初に引き込まれるポンプ作用である口腔ポンピングによって呼吸する。 その後、これらは閉じられ、空気は喉の収縮によって肺に強制される。 それらは湿った保たれる必要がある皮を通したガス交換とこれを補う。
カエルでは、骨盤ガードルは堅牢で、後肢は前肢よりもはるかに長く強くなります。 足は四、五桁を持っており、つま先は、多くの場合、水泳のために水かきをしたり、登山のための吸引パッドを持っています。 カエルは大きな目と尾を持っていません。 彼らの短い足は横に突出し、腹は地面に近いか、または接触しており、彼らは長い尾を持っています。 カイコは表面的にはミミズに似ており、手足がありません。 彼らは体に沿って移動し、彼らは左右に自分の体を起伏させることによって泳ぐ筋肉収縮のゾーンによって穴を掘る。H3>
主な記事
主な記事
主な記事
: 爬虫類の解剖学

爬虫類は、カメ、トゥアタラ、トカゲ、ヘビ、ワニ 彼らは四足動物ですが、ヘビやトカゲのいくつかの種は、手足を持っていないか、彼らの手足ははるかにサイズが縮小されています。 彼らの骨は骨化が良く、両生類の骨よりも骨格が強い。 歯は円錐形で、大部分が均一な大きさです。 表皮の表面の細胞は防水層を作成する角質のスケールに変更されます。 爬虫類は両生類がそうであるように呼吸のために彼らの皮膚を使用することができず、彼らの胸壁を拡大することによって彼らの肺に空気を引き 心臓は両生類のそれに似ていますが、酸素化された血流と脱酸素化された血流をより完全に分離する中隔があります。 生殖器系は、ほとんどの種に存在する交尾器官で、内部受精のために進化してきました。 卵は乾燥からそれらを防ぎ、土地に置かれるか、またはある種で内部的に成長する羊膜によって囲まれる。 膀胱は窒素性廃棄物が尿酸として排泄されるので小さい。
カメは彼らの保護殻のために注目すべきです。 彼らは上の角質の甲羅と下のplastronに包まれた柔軟性のない幹を持っています。 これらは角質の物によって覆われ、肋骨および脊柱と部分的に溶けている皮膚で埋め込まれる骨の多い版から形作られます。 首は長く、適用範囲が広く、頭部および足は貝の中で引き戻すことができる。 カメは菜食主義者であり、典型的な爬虫類の歯は鋭い角質のプレートに置き換えられています。 水生種では、前脚は足ひれに変更されています。
Tuatarasは表面的にトカゲに似ていますが、系統は三畳紀に分岐しました。 一つの生きている種、Sphenodon punctatusがあります。 頭蓋骨は両側に2つの開口部(窓)を有し、顎は頭蓋骨にしっかりと取り付けられている。 下顎には一列の歯があり、これは動物が噛むときに上顎の二列の間に収まる。 歯は顎からの骨の多い材料のただ投射で、結局身に着けている。 脳と心臓は他の爬虫類のものよりも原始的であり、肺は単一の部屋を持ち、気管支を欠いています。 トゥアタラは、その額によく発達した頭頂眼を持っています。
トカゲは、それぞれの側に一つの窓だけで頭蓋骨を持っており、第二の窓の下の骨の下のバーが失われています。 これは口がより広く開くようにするより少なく堅く付す顎で起因する。 トカゲは主に四足で、胴体は短く横向きの脚で地面から離されていますが、いくつかの種は四肢を持たず、ヘビに似ています。 トカゲは可動性の眼瞼を有し、鼓膜が存在し、いくつかの種は中央の頭頂眼を有する。
ヘビはトカゲと密接に関連しており、白亜紀の間に共通の祖先の系統から分岐しており、同じ特徴の多くを共有しています。 骨格は頭蓋骨、舌骨、脊椎および肋骨からなるが、いくつかの種は骨盤棘の形で骨盤および後肢の痕跡を保持する。 第二のフェネストラの下のバーも失われており、顎はヘビが獲物全体を飲み込むことを可能にする極端な柔軟性を持っています。 ヘビは可動性のまぶたを欠いており、目は透明な”光景”の鱗で覆われています。 彼らは鼓膜を持っていませんが、彼らの頭蓋骨の骨を通して地面の振動を検出することができます。 彼らの分岐した舌は味と香りの器官として使用され、いくつかの種は温血動物の獲物を見つけることを可能にする彼らの頭の上に感覚ピットを持ワニは、長い鼻と多数の歯を持つ大型の低投げ水生爬虫類です。
ワニは、長い鼻と多数の歯を持つ大型の低投げ水生爬虫類です。
ワニは、長い鼻と多数の 頭部と胴部は背腹側に平らになり、尾部は横方向に圧縮される。 それは泳ぐとき水を通して動物を強制するために左右にうねります。 頑丈な角質化した鱗はボディアーマーを提供し、いくつかは頭蓋骨に融合しています。 鼻孔、目および耳は平らな頭部の上で上がり動物が浮かんでいるとき水の表面の上に残ることをそれらが可能にする。 バルブは、それが水没したときに鼻孔と耳を密封します。 他の爬虫類とは異なり、ワニは、酸素化された血液と脱酸素化された血液の完全な分離を可能にする四つの部屋を持つ心を持っています。
鳥の解剖学編集

鳥は四肢動物ですが、後肢は歩行やホッピングに使用されますが、前肢は羽で覆われ、飛行に適応しています。 鳥は吸熱性であり、高い代謝率、軽い骨格系および強力な筋肉を有する。 長い骨は薄く、中空で非常に軽い。 肺からの空気嚢の拡張は、いくつかの骨の中心を占めています。 胸骨は広く、通常はキールがあり、尾椎は融合しています。 歯はなく、狭い顎は角で覆われたくちばしに適応しています。 目は比較的大きく、特にフクロウのような夜行性の種では大きい。 彼らは捕食者で前方に、アヒルでは横に直面しています。
羽は表皮の外生であり、皮膚の上に扇状に広がる局所的なバンドに見られます。 大きな飛行羽は翼と尾に見られ、輪郭の羽は鳥の表面を覆い、若い鳥や水鳥の輪郭の羽の下に細かい羽毛が発生します。 唯一の皮膚腺は、尾の基部近くの単一のuropygial腺である。 これは、鳥がpreensときに羽を防水油性分泌物を生成します。 つま先の先端には足、足、爪に鱗があります。
哺乳類の解剖学編集
哺乳類は、主に陸生動物の多様なクラスですが、いくつかは水生であり、他の人は羽ばたきや滑空飛行を進化させてきました。 彼らは主に四肢を持っていますが、いくつかの水生哺乳類は手足を持っていないか、手足がフィンに変更され、コウモリの前肢が翼に変更されています。 ほとんどの哺乳類の脚は胴体の下に位置しており、地面から十分に離れています。 哺乳類の骨はよく骨化されており、通常は区別される歯は角柱状のエナメル質の層で覆われています。 歯はクジラ類の場合のように、動物の生涯の間に一度(乳歯)流されるか、まったく流されません。 哺乳類は中耳に3つの骨と内耳に1つの蝸牛を持っています。 彼らは髪に服を着て、彼らの皮膚には汗を分泌する腺が含まれています。 これらの腺のいくつかは、若者を養うために牛乳を生産し、乳腺として特化しています。 哺乳類は肺で呼吸し、胸郭を腹部から分離する筋肉の横隔膜を持ち、肺に空気を引き込むのに役立ちます。 哺乳類の心臓には4つの室があり、酸素化された血液と脱酸素化された血液は完全に分離されています。 窒素性廃棄物は主に尿素として排泄される。
哺乳動物は羊水であり、ほとんどが胎生であり、若く生きることを出産する。 これの例外は、卵を産むmonotreme、カモノハシ、オーストラリアのエキドナです。 他のほとんどのほ乳類に成長の胎児が滋養物を得る胎盤がありますが、有袋類で、胎児の段階は非常に短く、未熟な若者は生まれ、ニップルに掛け金を
人間の解剖学編集

人間は哺乳類の全体的な体の計画を持っています。 人間は頭、首、胴(胸郭と腹部を含む)、2つの腕と手、そして2つの足と足を持っています。
一般的に、特定の生物科学の学生、救急医、義肢装具士、理学療法士、作業療法士、看護師、足病医、および医学生は、解剖学的モデル、骨格、教科書、図、写真、講義やチュー 顕微鏡解剖学(または組織学)の研究は、顕微鏡下で組織学的調製物(またはスライド)を調べる実践的な経験によって支援することができる。人間の解剖学、生理学、生化学は補完的な基本的な医学であり、一般的に医学部の最初の年に医学生に教えられています。
人間の解剖学、生理学、生化学は、 すなわち、それぞれ、頭部や胸部などの身体領域による解剖学の研究、または神経系や呼吸器系などの特定のシステムによる研究である。 主要な解剖学の教科書、グレイの解剖学は、現代の教授法に沿って、システム形式から地域形式に再編成されました。 解剖学の完全な実用的な知識は、医師、特に外科医および組織病理学および放射線学などのいくつかの診断専門分野で働く医師によって必要とされ
学術解剖学者は、通常、大学、医学部または教育病院で採用されています。 彼らはしばしば解剖学を教え、特定のシステム、器官、組織または細胞の研究に関与しています。